|
||
|
|
||
| Главная ≫ Инфотека ≫ Математика ≫ Идеи Алана Тьюринга помогли понять механизм развития пальцев у позвоночных // Александр Марков |
Идеи Алана Тьюринга помогли понять механизм развития пальцев у позвоночныхАлександр Марков
Генно-инженерные эксперименты показали, что количество пальцев у мышей зависит от двух взаимодействующих систем генов-регуляторов. По мере отключения этих генов пальцы становятся многочисленнее, короче и тоньше, а их концы соединяются костно-хрящевой дугой, так что в итоге кисть начинает напоминать плавник примитивной рыбы. Новые данные согласуются с гипотезой о том, что развитие пальцев основано на реакционно-диффузионном механизме самоорганизации, придуманном Аланом Тьюрингом в 1952 году. Этот механизм способен генерировать сложные узоры из полос и пятен, часто встречающиеся в живой природе. По-видимому, пальцы развиваются там, где образуются неподвижные «волны» концентраций двух динамически взаимодействующих веществ (морфогенов). Сами эти вещества пока не идентифицированы, а известные гены-регуляторы лишь модифицируют силу их взаимного воздействия, что отражается на длине «волн», которая, в свою очередь, определяет ширину пальцев и их количество. Алан Тьюринг (1912–1954) внес важный вклад не только в информатику, но и в теорию морфогенеза. Он предположил, что некоторые сложные, упорядоченные структуры могут формироваться путем самоорганизации на основе реакционно-диффузионного механизма, называемого также «моделью Тьюринга». Эта модель, детально разработанная математиками уже после преждевременной смерти Тьюринга, предполагает наличие двух динамически взаимодействующих веществ (морфогенов). Первое вещество, активатор, усиливает собственное производство (положительная обратная связь), а также производство второго вещества — ингибитора. Ингибитор подавляет активность или уменьшает количество активатора (отрицательная обратная связь) и самого себя. Оба вещества проникают путем диффузии из клетки в клетку, но делают это с разной скоростью. Такая система в зависимости от параметров (силы взаимных влияний и скоростей диффузии) спонтанно порождает разнообразные «рисунки» изменения концентраций в пространстве, которые могут быть как подвижными, вроде бегущих волн, так и стационарными. Предполагается, что реакционно-диффузионные механизмы лежат в основе формирования пятен на шкуре леопарда, полосок зебры и других замысловатых узоров и орнаментов (рис. 2; см. также: Джеймс Д. Марри. Отчего у леопарда пятна на шкуре).
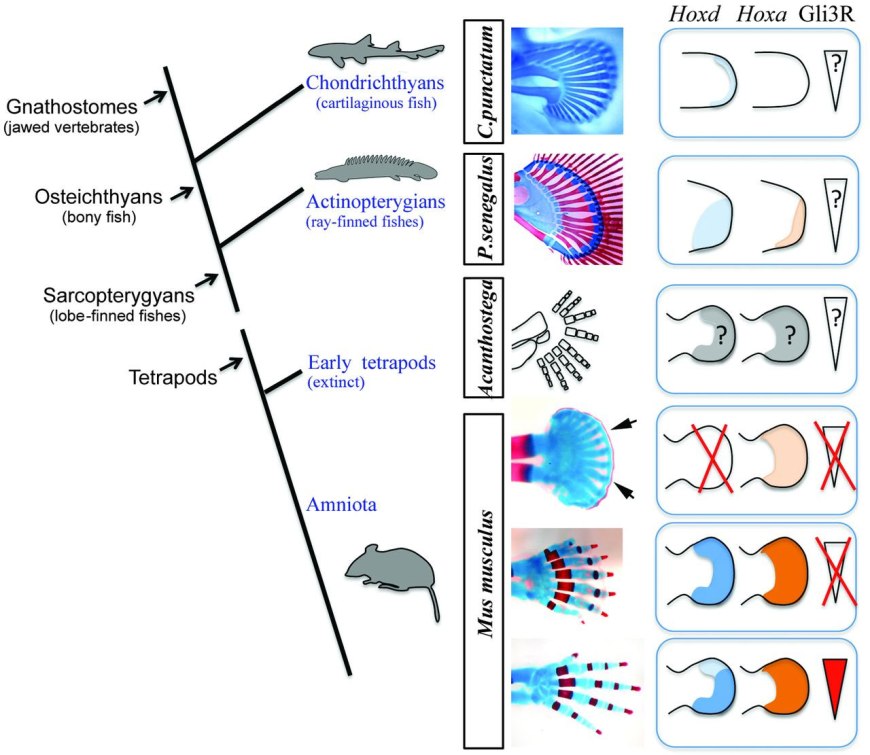
Новое исследование, выполненное биологами из Испании, Канады и США, показало, что реакционно-диффузионный механизм, возможно, отвечает и за формирование пальцев у позвоночных. В ходе онтогенеза пальцы действительно возникают в виде «узора» из регулярно расположенных полосок на еще не расчлененной пластинке — зачатке кисти или стопы. Поначалу эти полоски отличаются от соседних клеток только активностью некоторых генов (таких как Sox9), а потом в них начинает формироваться хрящевая ткань. Важную роль в формировании пальцев играет сигнальная система, основанная на белках Sonic hedgehog (Shh) и Gli3. У заднего края зачатка кисти (там, где будет мизинец) включается «организатор» — группа клеток, выделяющих сигнальный белок-морфоген Shh. Последний не позволяет белку Gli3 превратиться в Gli3R — белок-репрессор, подавляющий экспрессию некоторых других генов-регуляторов. В результате формируется передне-задний градиент концентрации Gli3R (максимум там, где будет большой палец, минимум — возле мизинца), который играет ключевую роль в определении передне-задней полярности зачатка конечности. Мутации, нарушающие работу системы Shh/Gli3, могут приводить к полидактилии (см.: Рыбьи гены помогли понять, почему первые четвероногие были многопалыми) и нарушениям «идентичности» пальцев (например, вместо большого пальца может развиться второй мизинец). Однако эта система, вопреки прежним предположениям, не является необходимой для развития пальцев как таковых. Оказалось, что даже если ее полностью отключить (например, удалив ген Gli3), пальцы всё равно развиваются (хоть и не такие, как надо, и в большем числе). Это навело некоторых эмбриологов на мысль, что формирование пальцев, возможно, основано не на «позиционной информации», задаваемой градиентом концентрации того или иного морфогена (как думали раньше), а на совсем другом принципе. Например, на тьюринговском реакционно-диффузионном механизме. Для проверки этого предположения авторы изучили эффекты другой важной группы генов-регуляторов, экспрессирующихся в зачатках кистей и стоп — хокс-генов Hoxa13, Hoxd11, Hoxd12 и Hoxd13. Их роль в развитии пальцев до сих пор оставалась не совсем ясной. Генно-инженерные эксперименты на мышах показали, что у животных с нормально работающей системой Shh/Gli3 частичное отключение перечисленных хокс-генов не приводит к выраженной полидактилии, однако ведет к укорочению пальцев, уменьшению числа фаланг, а в экстремальных случаях — к формированию костно-хрящевой дуги, соединяющей кончики пальцев (рис. 1, верхний ряд). Самые интересные результаты дало отключение хокс-генов у мышей с неработающей системой Shh/Gli3 (нижний ряд на рис. 1). Оказалось, что постепенное уменьшение «дозы» хокс-генов (то есть числа работоспособных аллелей) приводит к столь же постепенному усилению полидактилии. У мышей с отключенной системой Shh/Gli3, но с полным набором хокс-генов (Gli3–/–, Hoxa13+/+, Hoxd11-13+/+) на лапках развивается по 7–8 пальцев. Отключение половины копий хокс-генов (Gli3–/–, Hoxa13+/–, Hoxd11-13+/–) увеличивает число пальцев до 8–9. Если же отключить гены Hoxd полностью (Gli3–/–, Hoxa13+/–, Hoxd11-13–/–) , пальцев становится уже 13. При этом пальцы становятся короткими, тонкими, почти не расчлененными, а их концы сливаются в цельную костно-хрящевую дугу, окаймляющую кисть. Такая конечность больше похожа на плавники акул и примитивных лучеперых рыб, чем на лапы нормальных четвероногих (рис. 3).
По мнению исследователей, полученные результаты прекрасно согласуются с версией о реакционно-диффузионном механизме формирования пальцев. Авторы разработали тьюринговскую модель, которая воспроизводит экспериментально установленные факты не только в общих чертах, но и в мелких деталях. Например, при полном отключении Hoxa13 эмбрионы погибают до рождения, но зачатки конечностей у них успевают сформироваться. Это позволило установить, что пальцы у эмбрионов Gli3–/–, Hoxa13–/– имеют тенденцию ветвиться. Такие же ветвящиеся пальцы сгенерировала и модель Тьюринга, когда в нее ввели соответствующие параметры (рис. 4).
Предложенная авторами модель постулирует существование двух взаимодействующих морфогенов — активатора и ингибитора. Оба морфогена активируются активатором и ингибируются ингибитором, причем с разной силой. Это порождает на периферии зачатка конечности неподвижные «волны» концентраций морфогенов в форме полос, соответствующих будущим пальцам. Чтобы пальцы были ориентированы правильно, то есть росли от основания к периферии, необходимо, чтобы сила положительного влияния активатора на самого себя регулировалась расстоянием от основания конечности (проксимально-дистальным градиентом, который задается у позвоночных морфогеном FGF). Для объяснения большинства установленных фактов достаточно допустить, что сила воздействия активатора на ингибитор зависит от «дозы» генов Hoxa13 и Hoxd11-13. Чем меньше доза, тем сильнее воздействие. Отключение хокс-генов усиливает влияние активатора на ингибитор, что приводит к уменьшению длины «волн» (подобно тому, как период колебаний пружинного маятника уменьшается с ростом жесткости пружины). В результате пальцы становятся многочисленнее и тоньше. Наконец, чтобы объяснить ряд тонких деталей (таких как изменение ширины пальцев в проксимально-дистальном направлении, а также случаи их разветвления), пришлось еще предположить, что эффект хокс-генов модулируется проксимально-дистальным градиентом FGF.
Получившаяся модель весьма точно воспроизвела эффект уменьшения дозы хокс-генов у мышей Gli3–/– (рис. 5), включая то обстоятельство, что по мере увеличения числа пальцев уменьшается их длина. При полном отключении Hoxa13 и Hoxd11-13 число пальцев должно стремиться к бесконечности, а их размер — к нулю, что на практике будет означать просто-напросто отсутствие пальцев. Именно это и наблюдается как в модели, так и у трансгенных эмбрионов (которые в этом случае не доживают до рождения). Если выводы авторов верны, то искусственное усиление активности хокс-генов в зачатках конечности должно приводить к олигодактилии — уменьшению числа пальцев. Хорошо бы это проверить. Впрочем, отрицательный результат не опровергнет гипотезу, потому что не исключено, что нормальная активность хокс-генов уже обеспечивает максимальный возможный эффект. Решающей проверкой, конечно, будет обнаружение (или необнаружение после тщательных поисков) загадочных «тьюринговских» морфогенов — активатора и ингибитора. Сходство лапок трансгенных мышат с плавниками рыб (рис. 3) позволяет предположить, что тьюринговский механизм, контролирующий развитие конечностей, сформировался очень давно — возможно, еще до разделения челюстноротых позвоночных на хрящевых и костных рыб. Последние передали его четвероногим «по наследству». Возможно, превращение плавников в лапы было связано лишь с перенастройкой этого древнего механизма, с изменением характера влияния на него хокс-генов и Shh, а не с созданием принципиально новой программы развития конечностей. Источник: Rushikesh Sheth, Luciano Marcon, M. Félix Bastida, Marisa Junco, Laura Quintana, Randall Dahn, Marie Kmita, James Sharpe, Maria A. Ros. Hox Genes Regulate Digit Patterning by Controlling the Wavelength of a Turing-Type Mechanism // Science. 2012. V. 338. P. 1476–1980. См. также: Александр Марков
ТегиПохожее
|





 Возможно, в основе широкого разнообразия раскрасок шкуры у животных, наблюдаемого в природе, лежит единый механизм формирования таких структур. Результаты математического моделирования этого механизма открывают биологам новые перспективы для исследований.
Возможно, в основе широкого разнообразия раскрасок шкуры у животных, наблюдаемого в природе, лежит единый механизм формирования таких структур. Результаты математического моделирования этого механизма открывают биологам новые перспективы для исследований. В научных представлениях о происхождении жизни в последнее десятилетие происходит настоящая революция, и она далеко не завершена. К сожалению, эта информация доступна в основном только на английском языке. Цикл статей, предлагаемый вниманию читателей, отчасти восполнит этот пробел.
В научных представлениях о происхождении жизни в последнее десятилетие происходит настоящая революция, и она далеко не завершена. К сожалению, эта информация доступна в основном только на английском языке. Цикл статей, предлагаемый вниманию читателей, отчасти восполнит этот пробел. Продолжительность циклов большинства цикад не случайна, а представляет собой интервалы из простых чисел (чисел, делимых без остатка только на себя — 3, 5, 7, 11, 13, 17 и т. д.), являясь наиболее действенной стратегией выживания и размножения.
Продолжительность циклов большинства цикад не случайна, а представляет собой интервалы из простых чисел (чисел, делимых без остатка только на себя — 3, 5, 7, 11, 13, 17 и т. д.), являясь наиболее действенной стратегией выживания и размножения. Окраска многих животных устроена причудливо и замысловато. На клеточном уровне ее возникновение описывается реакционно-диффузными моделями при помощи систем дифференциальных уравнений. В недавней работе группа ученых из Швейцарии детально изучила механизм формирования окраски глазчатых ящериц Timon lepidus. Оказалось, что это происходит по правилам, характерным для дискретного клеточного автомата, где в роли ячеек автомата выступают отдельные чешуйки кожи ящериц. Математическое моделирование позволило понять, что реакционно-диффузная система может порождать клеточный автомат благодаря особым условиям — в данном случае это подходящие размеры чешуек и толщина кожи ящериц внутри и на границе чешуек.
Окраска многих животных устроена причудливо и замысловато. На клеточном уровне ее возникновение описывается реакционно-диффузными моделями при помощи систем дифференциальных уравнений. В недавней работе группа ученых из Швейцарии детально изучила механизм формирования окраски глазчатых ящериц Timon lepidus. Оказалось, что это происходит по правилам, характерным для дискретного клеточного автомата, где в роли ячеек автомата выступают отдельные чешуйки кожи ящериц. Математическое моделирование позволило понять, что реакционно-диффузная система может порождать клеточный автомат благодаря особым условиям — в данном случае это подходящие размеры чешуек и толщина кожи ящериц внутри и на границе чешуек. Закон квадрата — куба представляет собой следующий принцип: если объект пропорционально (то есть с помощью преобразования подобия) увеличивается (уменьшается) в размере, его новый объём будет пропорционален кубу масштабирующего коэффициента, а новая площадь его поверхности — пропорциональна квадрату масштабирующего коэффициент. Этот закон находит своё применение в технике и биомеханике.
Закон квадрата — куба представляет собой следующий принцип: если объект пропорционально (то есть с помощью преобразования подобия) увеличивается (уменьшается) в размере, его новый объём будет пропорционален кубу масштабирующего коэффициента, а новая площадь его поверхности — пропорциональна квадрату масштабирующего коэффициент. Этот закон находит своё применение в технике и биомеханике. Рассмотрены строение и механизмы действия протонной АТРсинтазы и флагеллярного мотора - молекулярных моторов живой клетки, выполняющих химическую и механическую работу, связанную с их вращательным движением, строение и механизмы работы миозина и кинезина - механохимических белков, ответственных за сократительную активность и внутриклеточный транспорт органелл в клетке.
Рассмотрены строение и механизмы действия протонной АТРсинтазы и флагеллярного мотора - молекулярных моторов живой клетки, выполняющих химическую и механическую работу, связанную с их вращательным движением, строение и механизмы работы миозина и кинезина - механохимических белков, ответственных за сократительную активность и внутриклеточный транспорт органелл в клетке. Иногда простая математическая модель хорошо описывает сложную биологическую систему. Примером этого служат долговременные отношения между видами хищника и жертвы в какой-либо экосистеме. Математические расчеты роста популяции отдельно взятого вида показывают, что пределы плотности популяции можно описать простыми уравнениями, которые на выходе дают характерную S-образную кривую. Это — кривая численности популяции, которая растет экспоненциально, пока она небольшая, а затем выравнивается, когда она достигает пределов возможности экосистемы поддерживать ее. Простое продолжение этой концепции позволяет нам понять экосистему, в которой взаимодействуют два вида — хищник и жертва.
Иногда простая математическая модель хорошо описывает сложную биологическую систему. Примером этого служат долговременные отношения между видами хищника и жертвы в какой-либо экосистеме. Математические расчеты роста популяции отдельно взятого вида показывают, что пределы плотности популяции можно описать простыми уравнениями, которые на выходе дают характерную S-образную кривую. Это — кривая численности популяции, которая растет экспоненциально, пока она небольшая, а затем выравнивается, когда она достигает пределов возможности экосистемы поддерживать ее. Простое продолжение этой концепции позволяет нам понять экосистему, в которой взаимодействуют два вида — хищник и жертва. Почти всякая незыблемая общепринятая теория, которую с проклятьями зубрят школьники и которую устало и одинаково рассказывают учителя и даже профессора ВУЗов, при внимательном рассмотрении оказывается отнюдь не однозначной, захватывающей и полной загадок. К теории мышечного сокращения вышесказанное относится в полной мере. В общих чертах она была разработана еще в 50-х годах прошлого века, и классический рисунок с актиновыми и миозиновыми нитями до сих пор кочует из учебника в учебник. Однако реальная картина сокращения мышцы куда запутаннее, интереснее и непонятнее, со множеством подробностей и неожиданных действующих лиц и со сложными ролями, которые исполняют эти лица.
Почти всякая незыблемая общепринятая теория, которую с проклятьями зубрят школьники и которую устало и одинаково рассказывают учителя и даже профессора ВУЗов, при внимательном рассмотрении оказывается отнюдь не однозначной, захватывающей и полной загадок. К теории мышечного сокращения вышесказанное относится в полной мере. В общих чертах она была разработана еще в 50-х годах прошлого века, и классический рисунок с актиновыми и миозиновыми нитями до сих пор кочует из учебника в учебник. Однако реальная картина сокращения мышцы куда запутаннее, интереснее и непонятнее, со множеством подробностей и неожиданных действующих лиц и со сложными ролями, которые исполняют эти лица.| Главная ≫ Инфотека ≫ Математика ≫ Идеи Алана Тьюринга помогли понять механизм развития пальцев у позвоночных // Александр Марков |
|
[time: 10 ms; queries: 7]
19 Апр 2024 14:41:08 GMT+3 |